- 移动端


上海百趣生物科技有限公司
13 年
手机商铺
商家活跃:
产品热度:
- NaN
- 0.7000000000000002
- 0.7000000000000002
- 2.7
- 2.7
植物阿趣广靶®代谢组学
询价
推荐产品





公司新闻/正文
PLANT CELL ENVIRON|中国科学院昆明植物研究所解密大花蕙兰花色的时空调控机制
103 人阅读发布时间:2025-04-07 17:08

文章标题:Spatial and Temporal Regulation of Flower Coloration in Cymbidium lowianum
中文标题:大花蕙兰花色的时空调控
发表期刊:Plant, Cell & Environment
影响因子:6.5
研究背景
花色是吸引传粉者并决定花卉观赏价值的关键性状。大花蕙兰(Cymbidium lowianum)是兰属杂交种的重要亲本之一,具有两种花色形态(正常花和白化花),其唇瓣颜色存在差异。然而,目前对大花蕙兰花色形成的分子机制尚不清楚。本研究通过比较正常花和白化花唇瓣组织的代谢组分析,发现矢车菊素-3-O-葡萄糖苷含量在红色唇瓣中显著高于其他唇瓣组织,表明该化合物是大花蕙兰花色变异和唇瓣颜色分化的原因(图1 A-B)。此外,研究还发现红色花色与F3'H基因表达水平呈正相关,且花发育过程中,花青素、类胡萝卜素和叶绿素共同影响萼片和花瓣的颜色变化。
研究结果
1.花色相关代谢物和基因表达差异
本研究通过代谢组学分析,比较了正常花和白化花唇瓣组织中的代谢物差异。结果显示,正常花的红色唇瓣(epichile)中矢车菊素-3-O-葡萄糖苷(cyanidin-3-O-glucoside)含量显著高于其他唇瓣组织(图1C),表明其是导致花色差异的关键花青素。同时,白化花的唇瓣中二氢杨梅素(dihydromyricetin)、黄酮醇(flavonols)、山柰酚(kaempferol)和槲皮素(quercetin)含量较高。这些代谢物的差异表明,花色的形成与花青素的种类和浓度密切相关。
在基因表达层面,通过RNA测序分析发现,与花青素生物合成途径相关的结构基因在正常花的唇瓣中表达水平较高,尤其是F3'H基因在正常花的红色唇瓣中表达量最高(图1D-E)。这进一步证实了F3'H基因在花色形成中的关键作用。
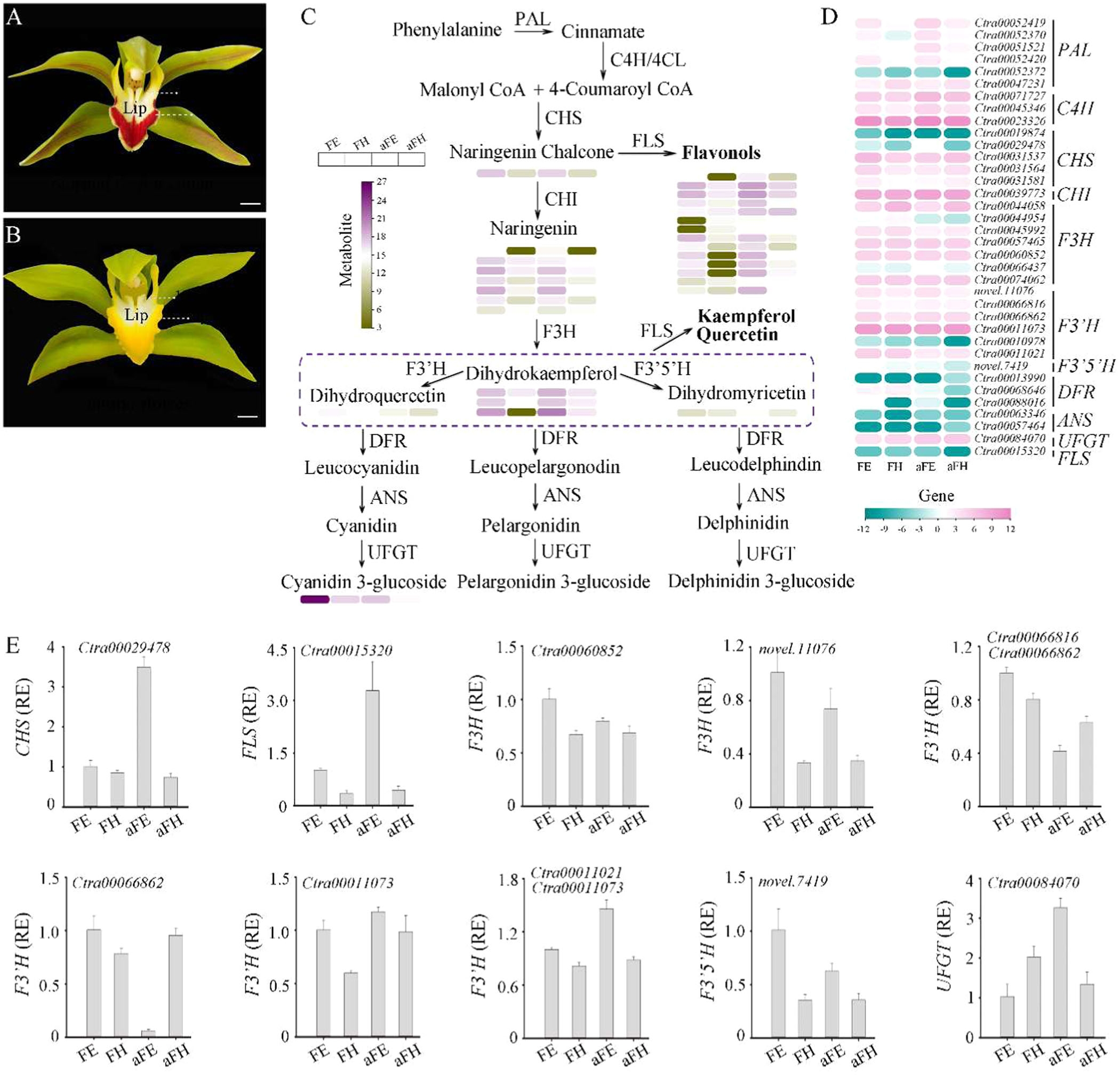
图1.大花蕙兰花青素生物合成途径关键代谢产物及结构基因综述
2.转录因子的差异表达
研究进一步分析了正常花和白化花唇瓣中转录因子的表达差异。通过转录组数据,鉴定出258个转录因子,包括bHLH、MYB和MADS-box家族成员。这些转录因子在花色形成中起着关键调控作用。通过对这些转录因子的表达模式进行聚类分析,发现某些转录因子在正常花的红色唇瓣中显著上调,如MYB14-1(图2D),而另一些则在白化花的唇瓣中上调,例如ODORANT1(图2E-F)。这些结果表明,这些转录因子可能通过调控花青素生物合成途径中的结构基因,进而影响花色的形成。
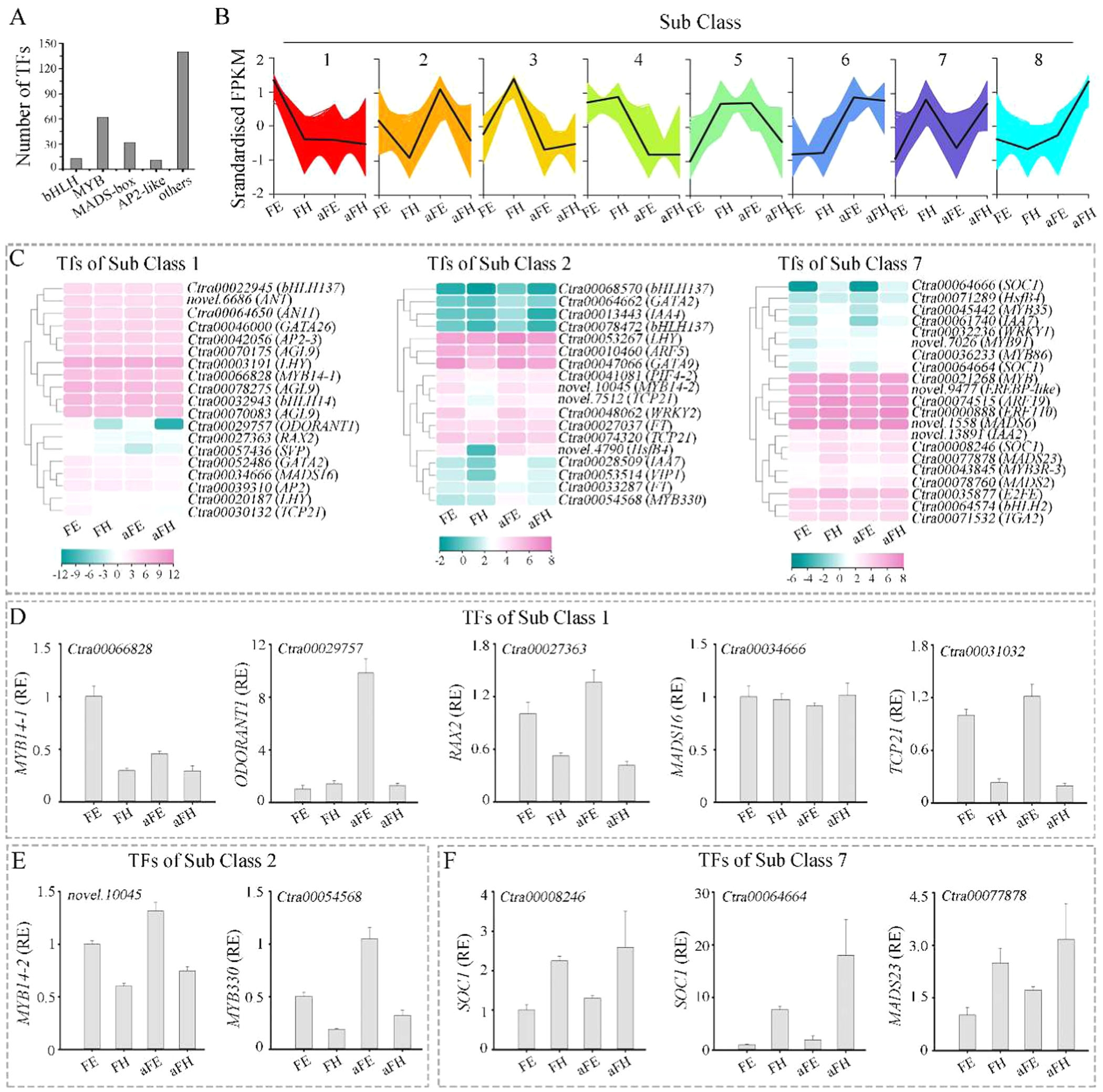
图2.花唇色形成的基因调控
3.花瓣和萼片发育过程中的色素积累和基因富集模式
大花蕙兰的花色在花蕾阶段到开花阶段会发生显著变化。研究者对花瓣和萼片在花蕾阶段、开花后5天(F5)和开花后10天(F10)的花青素、类胡萝卜素和叶绿素含量进行了定量分析。结果显示,从花蕾阶段到开花阶段,花瓣和萼片中的花青素和类胡萝卜素含量增加,而叶绿素含量则逐渐减少(图3C-E)。这种色素含量的变化与花色从绿色到深黄色的转变相一致。此外,矢车菊素-3-O-葡萄糖苷的含量在花发育过程中显著增加(图3F),进一步证实了其在花色变化中的重要作用。
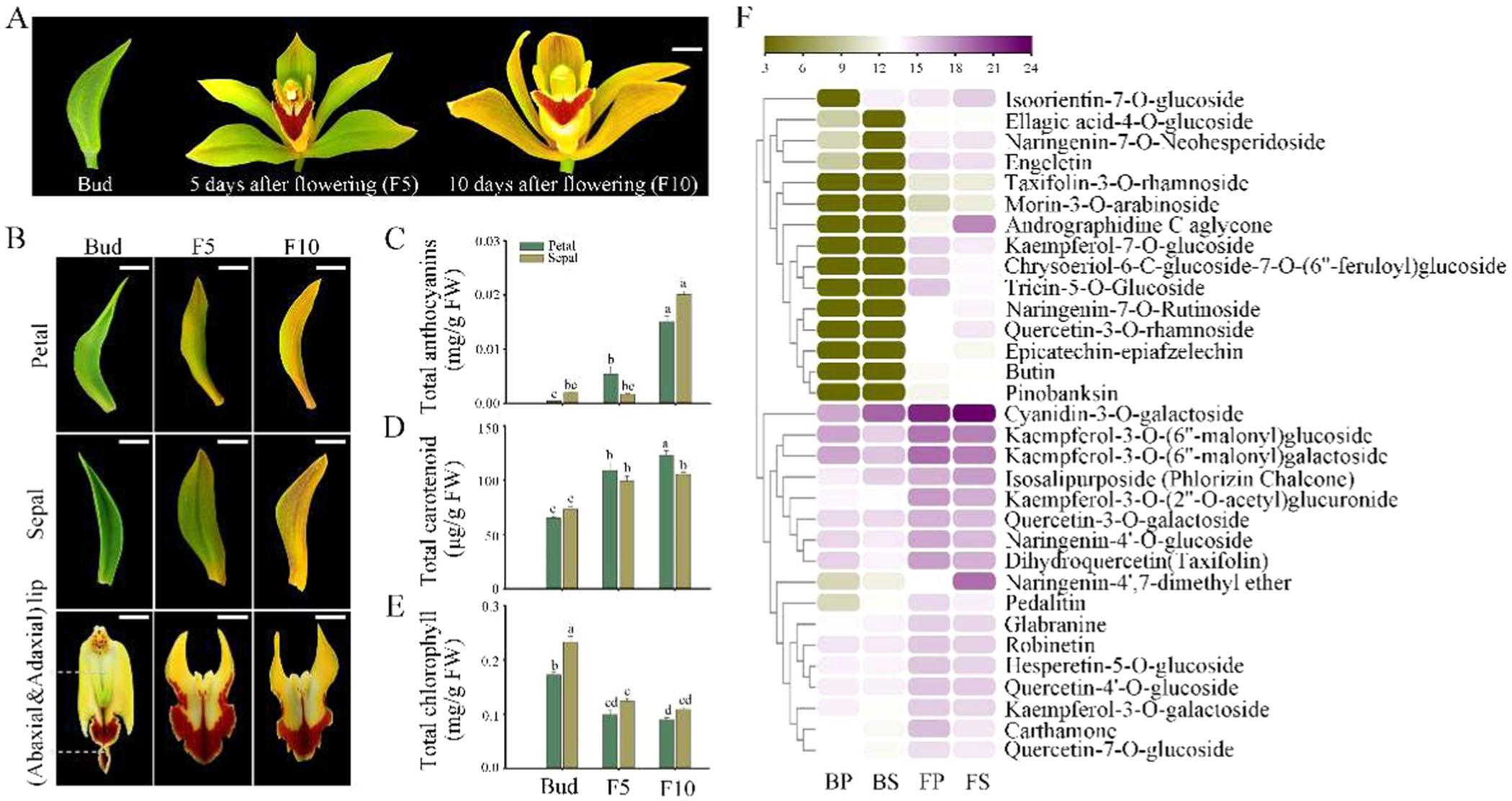
图3.正常花在花发育过程中关键色素的定量分析
转录组分析还揭示了花发育过程中差异表达基因(DEGs)的模式。研究发现,在花瓣和萼片中共有2711个基因差异表达,其中1663个基因仅在花瓣中差异表达,849个基因仅在萼片中差异表达(图4B)。这些差异表达基因主要富集在植物激素信号转导和萜类生物合成途径中(图4C)。这些结果表明,植物激素和萜类化合物可能在花色变化中起重要作用。
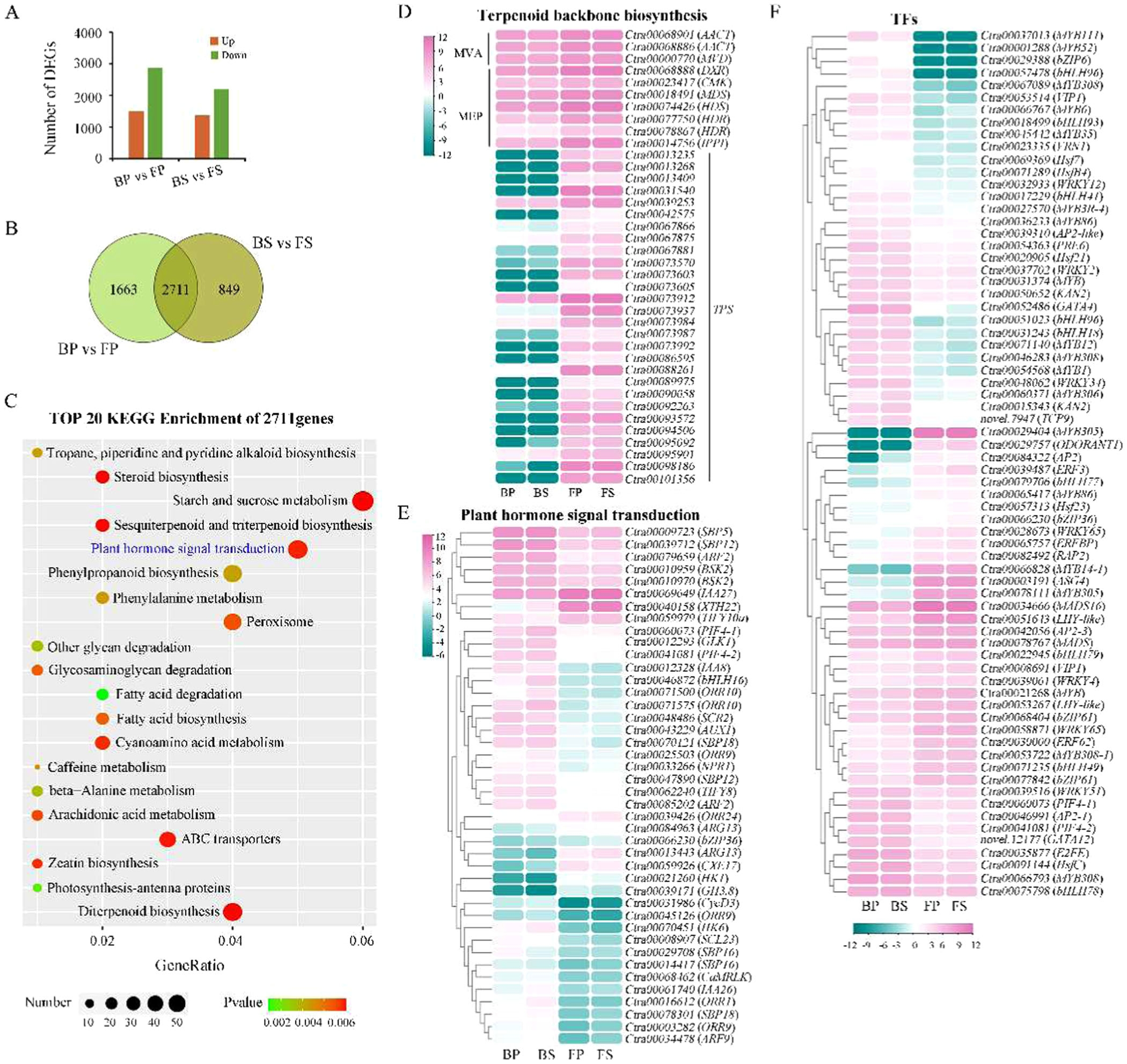
图4.大花惠兰花朵发育过程中差异表达基因(DEGs)的转录组分析
4.与色素生物合成相关的关键结构基因的表达分析
为了深入理解花色形成的分子机制,研究者筛选了与花青素、类胡萝卜素和叶绿素生物合成相关的关键结构基因,并分析了它们在花发育过程中的表达模式。结果显示,大多数与花青素和类胡萝卜素生物合成相关的基因在花发育过程中上调,而与叶绿素生物合成相关的基因则下调(图5A-C)。例如,F3'H和BCH基因在花发育过程中表达量增加,而HEMG和CHLI基因表达量减少。
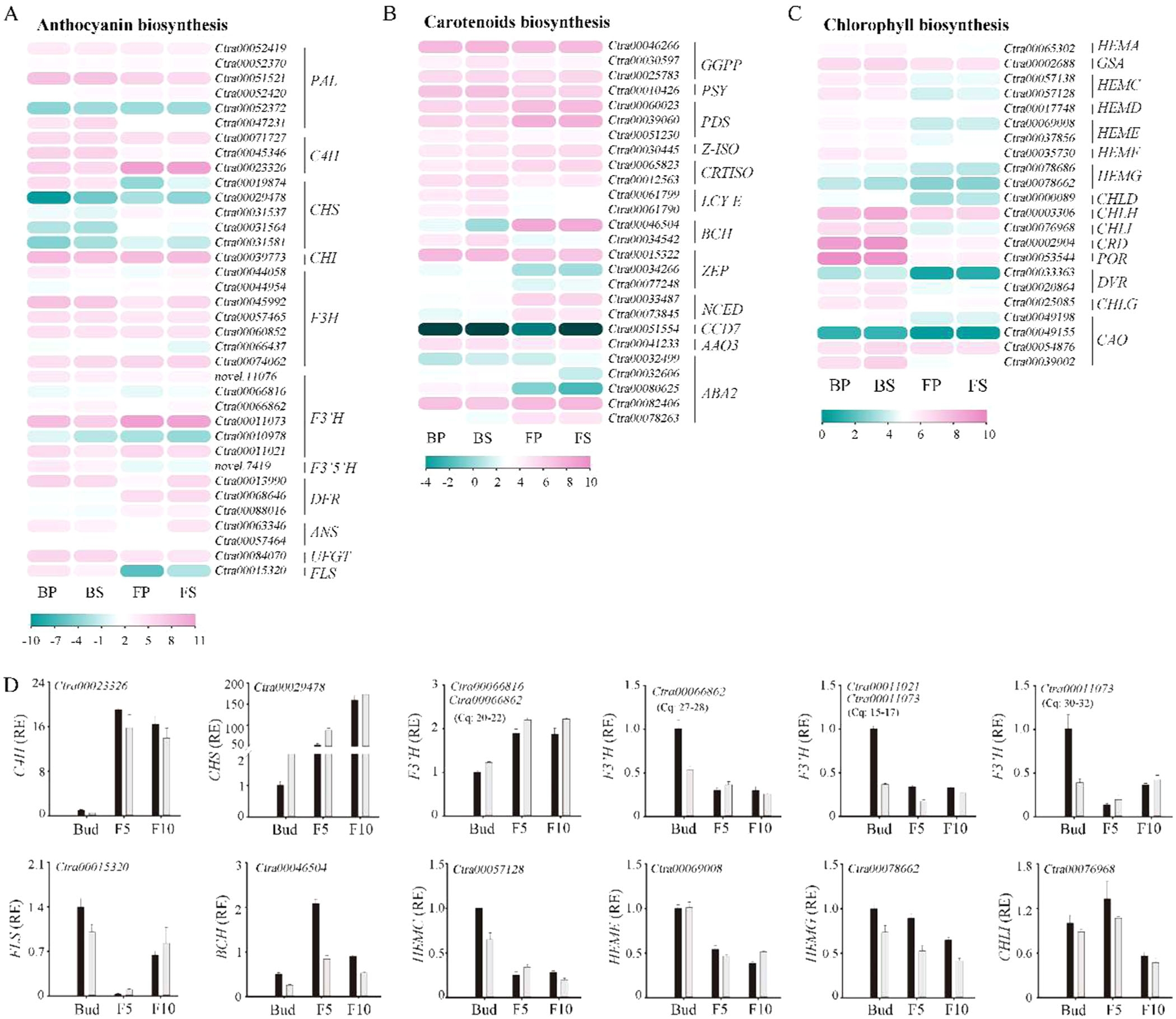
图5.色素生物合成相关结构基因的表达谱
为了验证这些结构基因的功能,研究者在拟南芥中构建了过表达转基因株系。结果表明,过表达F3'H的转基因拟南芥中花青素浓度显著增加,过表达BCH的转基因拟南芥中类胡萝卜素浓度增加,而过表达HEMG和CHLI的转基因拟南芥中叶绿素浓度增加(图6B-D)。这些结果进一步证实了这些结构基因在花色形成中的关键作用。
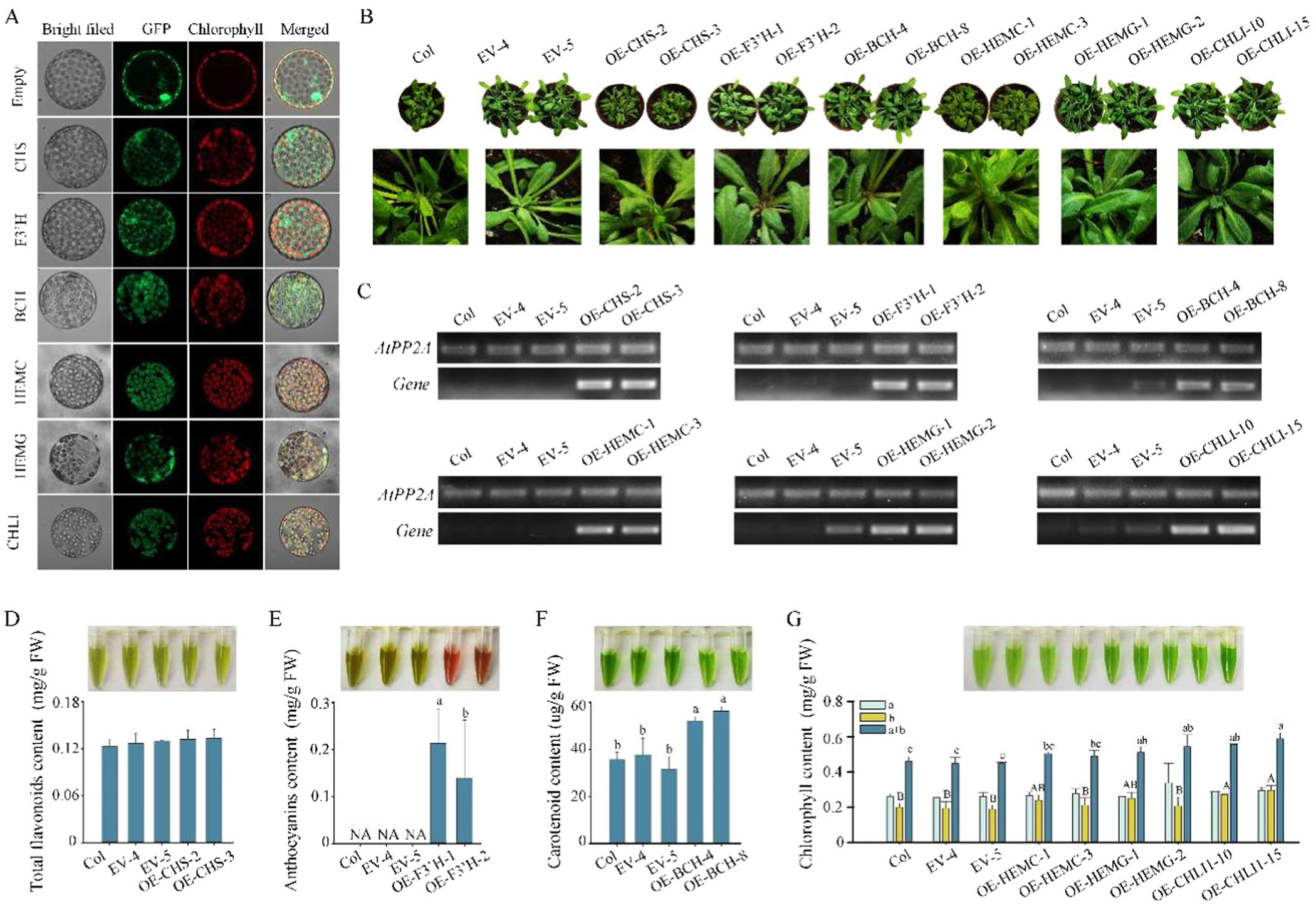
图6.参与色素生物合成的hub结构基因的功能分析
5.与色素生物合成相关的转录因子的鉴定
通过加权基因共表达网络分析(WGCNA) ,研究者鉴定出与花青素、类胡萝卜素和叶绿素生物合成相关的转录因子模块。这些模块中的转录因子包括MYB、bHLH和PIF家族成员(图7A-C)。例如,MYB14-1、MYB308-1和PIF4-2等转录因子与花青素和叶绿素的生物合成密切相关。
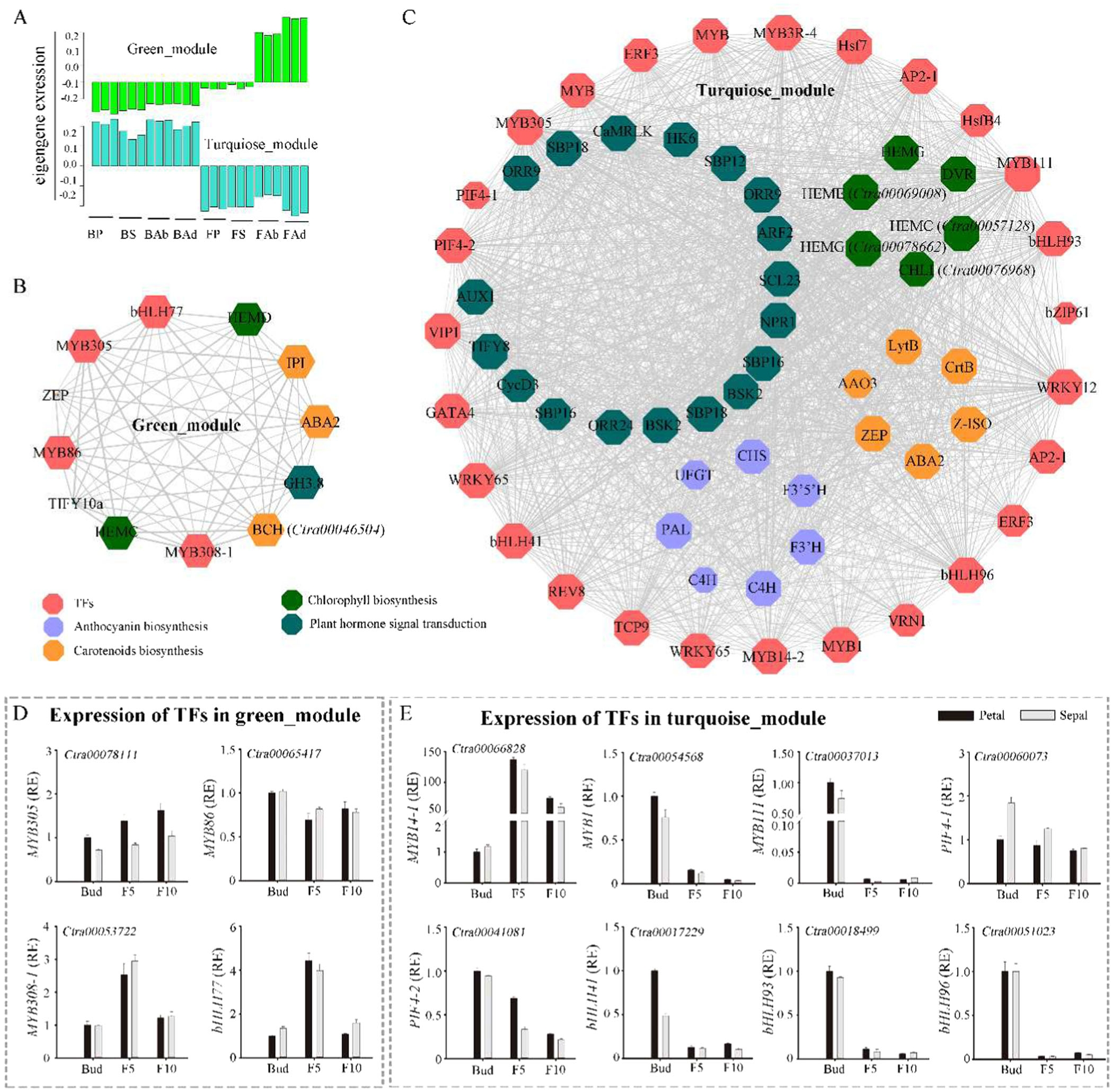
图7.黄姜色素生物合成的调控网络
为了验证这些转录因子的功能,研究者通过双荧光素酶报告基因实验和酵母单杂交实验,发现MYB14-1能够激活F3'H基因的表达,从而促进花青素的积累(图8B-E)。此外,PIF4-2与MYB14-1相互作用,共同调控花青素和叶绿素的生物合成(图9A)。这些结果表明,这些转录因子通过调控关键结构基因的表达,进而影响花色的形成。
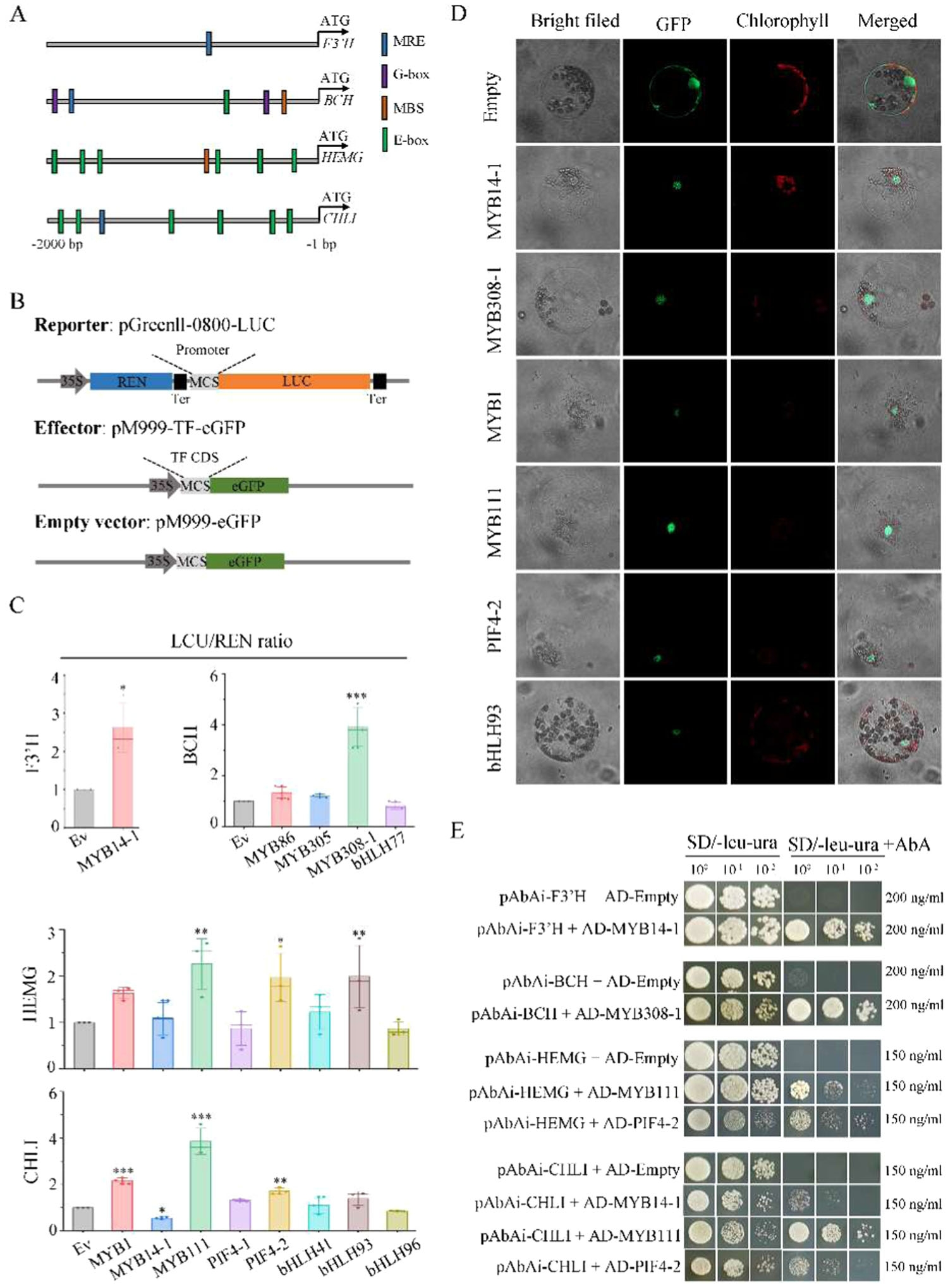
图8.参与激活色素生物合成结构基因表达的调控因子功能分析
6.PIF4-2与MYB14-1的相互作用
研究者通过酵母双杂交实验进一步验证了PIF4-2与MYB14-1之间的相互作用(图9A)。这种相互作用可能在花色调控中起到关键作用。PIF4-2作为光信号转导的关键因子,可能通过与MYB14-1的相互作用,调节花青素和叶绿素的生物合成,从而影响花色的形成。
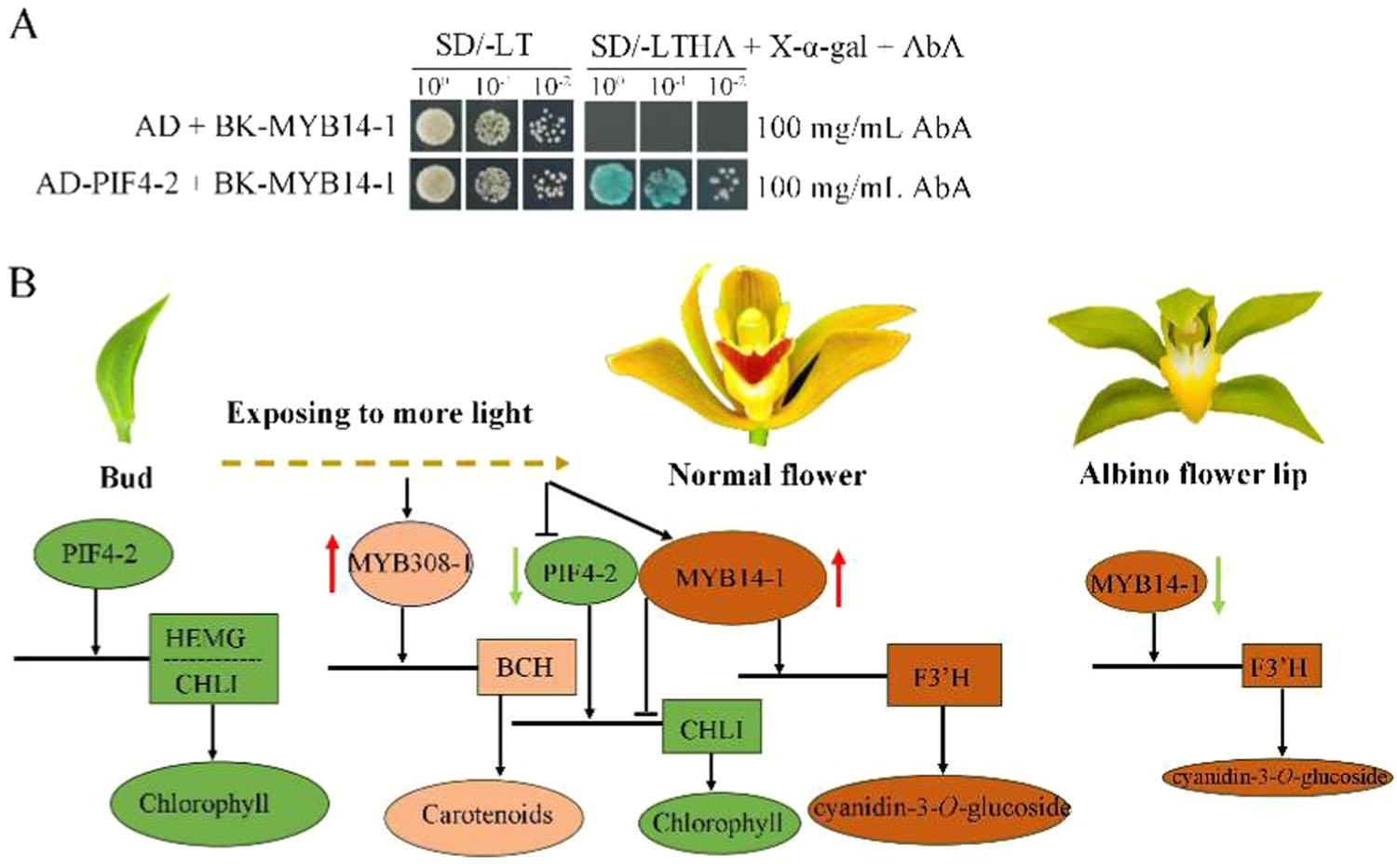
图9.PIF4‐2与MYB14‐1在大花惠兰花朵发育中的相互作用
研究结论
本研究揭示了大花蕙兰花色形成的分子机制,表明花色变异和唇瓣颜色分化主要由黄酮类化合物的类型和浓度决定,同时类胡萝卜素和叶绿素也在花发育过程中对花色产生影响。研究还鉴定了一系列关键的结构基因和转录因子,这些基因和转录因子通过调节花青素、类胡萝卜素和叶绿素的生物合成来影响花色。特别是,F3'H基因在花色形成中起关键作用,而PIF4-2与MYB14-1的相互作用可能是花色调控的重要机制。这些发现为兰花育种提供了重要的理论基础。